Perkembangan tumbuhan
Struktur penting dalam perkembangan tumbuhan adalah tunas, pucuk, akar, daun, dan bunga ; tumbuhan menghasilkan jaringan dan struktur ini sepanjang hidupnya dari meristem [1] yang terletak di ujung organ, atau di antara jaringan dewasa. Dengan demikian, tumbuhan hidup selalu memiliki jaringan embrionik. Sebaliknya, embrio hewan akan menghasilkan semua bagian tubuh yang akan dimilikinya sepanjang hidupnya sejak dini. Ketika hewan lahir (atau menetas dari telurnya), ia memiliki semua bagian tubuhnya dan sejak saat itu hanya akan tumbuh lebih besar dan lebih dewasa. Namun, baik tumbuhan maupun hewan melewati tahap filotipik yang berevolusi secara independen [2] dan yang menyebabkan kendala perkembangan yang membatasi penganekaan morfologis. [3] [4] [5] [6]
Menurut ahli fisiologi tumbuhan A. Carl Leopold, sifat-sifat organisasi yang terlihat pada tumbuhan adalah sifat-sifat munculan yang lebih dari sekadar jumlah bagian-bagian individu. "Perakitan jaringan dan fungsi-fungsi ini menjadi organisme multiseluler terpadu tidak hanya menghasilkan karakteristik bagian dan proses yang terpisah tetapi juga serangkaian ciri-cirinya baru yang tidak dapat diprediksi berdasarkan pemeriksaan bagian-bagian yang terpisah." [7]
Pertumbuhan
Tumbuhan berpembuluh dimulai dari zigot bersel tunggal, yang terbentuk melalui pembuahan sel telur oleh sel sperma. Dari titik itu, zigot mulai membelah untuk membentuk embrio tumbuhan melalui proses embriogenesis . Selama proses ini, sel-sel yang dihasilkan akan tersusun sedemikian rupa sehingga satu ujung menjadi akar pertama sementara ujung lainnya membentuk ujung tunas. Pada tumbuhan berbiji, embrio akan mengembangkan satu atau lebih "daun biji" ( kotiledon ). Pada akhir embriogenesis, tumbuhan muda akan memiliki semua bagian yang diperlukan untuk memulai kehidupannya.
Organogenesis tumbuhan
Setelah embrio berkecambah dari biji atau tanaman induknya, ia mulai menghasilkan organ tambahan (daun, batang, dan akar) melalui proses organogenesis . Akar baru tumbuh dari meristem akar yang terletak di ujung akar, dan batang serta daun baru tumbuh dari meristem tunas yang terletak di ujung tunas. [8] Percabangan terjadi ketika gumpalan kecil sel yang tertinggal oleh meristem, dan yang belum mengalami diferensiasi seluler untuk membentuk jaringan khusus, mulai tumbuh sebagai ujung akar atau tunas baru. Pertumbuhan dari meristem tersebut di ujung akar atau tunas disebut pertumbuhan primer dan menghasilkan pemanjangan akar atau tunas tersebut. Pertumbuhan sekunder menghasilkan pelebaran akar atau tunas dari pembelahan sel di kambium . [9]
Organogenesis langsung
Organogenesis langsung adalah metode budidaya jaringan tanaman di mana organ seperti akar dan tunas berkembang langsung dari sel meristematik atau non-meristematik, melewati tahap pembentukan kalus . Proses ini terjadi melalui penggiatan meristem apikal tunas dan akar atau tunas aksila, yang dipengaruhi oleh zat pengatur pertumbuhan tanaman yang diberikan secara internal atau eksternal. Akibatnya, tipe sel tertentu berdiferensiasi untuk membentuk struktur tanaman yang dapat tumbuh menjadi tanaman utuh. Teknik ini umumnya digunakan untuk memperbanyak berbagai spesies tanaman, termasuk sayuran, buah-buahan, tanaman berkayu, dan tanaman obat . Ujung tunas dan segmen nodus biasanya digunakan sebagai eksplan dalam proses ini. Dalam beberapa kasus, struktur adventif muncul dari jaringan somatik dalam kondisi tertentu, memungkinkan regenerasi tunas atau akar di area di mana mereka tidak akan berkembang secara alami. Pendekatan ini sangat efektif pada spesies herba, dan meskipun regenerasi adventif dapat menyebabkan tingkat pembentukan tunas yang lebih tinggi, proliferasi tunas aksila tetap menjadi metode yang paling banyak digunakan dalam mikropropagasi karena efisiensi dan kepraktisannya. Urutan umum perkembangan organ dalam proses ini mengikuti pola: Eksplan Primer → Meristemoid → Primordium Organ.
Organogenesis taklangsung
Organogenesis taklangsung adalah proses perkembangan di mana sel-sel tumbuhan mengalami dediferensiasi, memungkinkan mereka untuk kembali dari keadaan terspesialisasi dan bertransisi ke jalur perkembangan baru. Proses ini ditandai dengan tahap kalus perantara, di mana sel-sel kehilangan identitas aslinya dan menjadi mudah beradaptasi secara morfologis, yang berfungsi sebagai dasar pembentukan organ. Perkembangan organogenesis tidak langsung melibatkan beberapa fase kunci, dimulai dengan dediferensiasi, yang memungkinkan sel-sel untuk mencapai kompetensi, diikuti oleh tahap induksi yang mengarah ke keadaan yang sepenuhnya ditentukan. Setelah penentuan tercapai, sel-sel mengalami perubahan morfologis, yang pada akhirnya menghasilkan tunas atau akar yang fungsional. Proses ini mengikuti urutan perkembangan yang terstruktur: Eksplan Primer → Kalus → Meristemoid → Primordium Organ, memastikan pembentukan organ tumbuhan yang terorganisir.
Faktor pengaruh organogenesis
Eksplan
Kemampuan untuk meregenerasi tanaman secara sukses bergantung pada pemilihan eksplan yang tepat, yang bervariasi antar spesies dan varietas tanaman. Dalam organogenesis langsung, eksplan yang berasal dari jaringan meristematik, seperti ujung pucuk, tunas melintang, daun, tangkai daun, akar, dan struktur bunga, seringkali lebih disukai karena kemampuannya untuk berkembang dengan cepat menjadi organ baru. Jaringan-jaringan ini memiliki tingkat kelangsungan hidup yang tinggi, pertumbuhan yang cepat, dan potensi regenerasi yang kuat secara in vitro. Meristem, ujung pucuk, tunas aksilar, daun muda, dan embrio sangat efektif dalam mendorong regenerasi pada berbagai spesies tanaman.
Selain itu, bagian tanaman yang sudah dewasa, termasuk daun, batang, akar, tangkai daun, dan segmen bunga, juga dapat berfungsi sebagai eksplan yang layak untuk pembentukan organ dalam kondisi yang sesuai. Regenerasi tanaman terjadi melalui pembentukan kalus, massa sel yang belum terdiferensiasi yang kemudian menghasilkan organ baru. Pembentukan kalus dapat diinduksi dari berbagai eksplan, seperti kotiledon, hipokotil, batang, daun, pucuk apikal, akar, perbungaan, dan struktur bunga, ketika dikultur dalam kondisi terkontrol.
Secara umum, eksplan yang mengandung sel-sel yang aktif membelah lebih efektif untuk inisiasi kalus, karena memiliki kapasitas pemrograman ulang seluler yang lebih tinggi. Jaringan yang belum matang cenderung lebih mudah beradaptasi untuk regenerasi dibandingkan dengan jaringan yang matang karena plastisitas perkembangannya yang lebih tinggi. Ukuran dan bentuk eksplan juga memengaruhi keberhasilan pembentukan kultur, karena eksplan yang lebih besar atau lebih menguntungkan secara struktural dapat meningkatkan peluang bertahan hidup dan pertumbuhan. Perkembangan kalus terutama dipicu oleh luka dan keberadaan hormon tumbuhan, yang mungkin secara alami ada di dalam jaringan atau ditambahkan ke media pertumbuhan untuk merangsang aktivitas seluler dan pembentukan organ.
Media kultur, zat atur-tumbuh, dan agen gel.
Komposisi media kultur sangat bervariasi dalam kandungan unsur mineral dan vitaminnya untuk mengakomodasi kebutuhan beragam spesies tanaman. Media Murashige dan Skoog (MS) dibedakan oleh kandungan nitrogennya yang tinggi dalam bentuk amonium, karakteristik yang tidak ditemukan pada formulasi lain. Sukrosa biasanya berfungsi sebagai sumber karbohidrat utama di berbagai jenis media.
Interaksi antara auksin dan sitokinin dalam mengatur organogenesis sudah mapan, meskipun responsnya bervariasi menurut spesies. Beberapa tanaman, seperti tembakau, dapat secara spontan membentuk tunas tanpa regulator pertumbuhan eksogen, sementara yang lain seperti Scurrula pulverulenta, Lactuca sativa (Sawi India), dan Brassica juncea (Sawi India) sangat membutuhkan suplementasi hormonal. Dalam kultur kotiledon B. juncea, benzilaminopurin (BAP) saja menginduksi pembentukan tunas dari jaringan tangkai daun, mirip dengan pinus radiata di mana sitokinin saja sudah cukup untuk induksi tunas.[ diperlukan kutipan ]
Penelitian menunjukkan bahwa konsentrasi hormon endogen, bukan tingkat aplikasi eksogen, pada akhirnya menentukan diferensiasi organogenik. Di antara berbagai sitokinin (2iP, BAP, tidiazuron, kinetin, dan zeatin) yang digunakan untuk induksi tunas, BAP telah menunjukkan kemanjuran yang unggul dan penerapan yang luas. Auksin juga memengaruhi jalur organogenik, dengan 2,4-D umumnya digunakan untuk induksi kalus pada serealia, kendati organogenesis biasanya memerlukan transfer ke media yang mengandung IAA atau NAA atau sama sekali tidak mengandung 2,4-D. Rasio auksin terhadap sitokinin sebagian besar menentukan organ mana yang berkembang.
Asam giberelat (GA3) bersumbangsih pada pemanjangan sel dan pembentukan meristemoid, sementara senyawa non-konvensional seperti asam tri-iodobenzoat (TIBA), asam absisat (ABA), kanamisin, dan penghambat auksin telah terbukti efektif untuk spesies yang sulit diregenerasi. Aditif alami seperti bubuk ginseng dapat meningkatkan frekuensi regenerasi pada kultur tertentu. Karena etilen biasanya menekan diferensiasi tunas, penghambat sintesis etilen seperti aminoetoksivinilglisin (AVG) dan perak nitrat ( AgNO3 ) sering digunakan untuk mendorong organogenesis, dengan keberhasilan yang terdokumentasi pada kultur gandum, tembakau, dan bunga matahari.
Agar bukanlah komponen esensial dari media kultur, tetapi kualitas dan kuantitas agar merupakan faktor penting yang dapat menentukan perannya dalam organogenesis. Agar yang tersedia secara komersial mungkin mengandung pengotor. Dengan konsentrasi agar yang tinggi, media nutrisi menjadi keras dan tidak memungkinkan difusi nutrisi ke jaringan yang tumbuh. Hal ini memengaruhi proses organogenesis dengan menghasilkan akar adventif, kalus yang tidak diinginkan di pangkal, atau penuaan daun. pH adalah faktor penting lain yang dapat memengaruhi jalur organogenesis. pH media kultur disesuaikan antara 5,6 dan 5,8 sebelum sterilisasi. pH media memfasilitasi atau menghambat ketersediaan nutrisi dalam media; misalnya, penyerapan amonium in vitro terjadi pada pH stabil 5,5 (Thorpe et al., 2008).
Faktor-faktor lainnya
Musim dalam setahun
Waktu pengumpulan eksplan sangat memengaruhi kapasitas regeneratif dalam sistem kultur jaringan, dengan variasi musiman memainkan peran penting dalam keberhasilan pembentukan organ. Fenomena ini jelas ditunjukkan pada Lilium speciosum, di mana sisik umbi menunjukkan respons regeneratif yang berbeda berdasarkan musim pengumpulan. Eksplan yang dipanen selama musim semi dan musim gugur dengan mudah membentuk umbi kecil secara in vitro, sedangkan yang dikumpulkan selama bulan-bulan musim panas atau musim dingin gagal menghasilkan umbi kecil meskipun kondisi kultur identik.
Ketergantungan musiman serupa diamati pada Chlorophytum borivillianum, spesies yang bernilai obat yang menunjukkan peningkatan pembentukan umbi in vitro yang nyata selama musim hujan dibandingkan dengan waktu lain dalam setahun. Variasi musiman dalam potensi morfogenik ini kemungkinan mencerminkan perbedaan dalam keadaan fisiologis tanaman sumber, termasuk kadar hormon endogen, cadangan karbohidrat, dan aktivitas metabolisme yang berfluktuasi sepanjang siklus pertumbuhan tahunan.
Oksigen memiliki peran kunci dalam kultur jaringan, yang memengaruhi pembentukan organ. Dalam beberapa kultur, pembentukan tunas pucuk terjadi ketika gradien oksigen yang tersedia di dalam wadah kultur berkurang, sedangkan induksi akar membutuhkan gradien oksigen yang tinggi.
Lampu
Kondisi cahaya, termasuk intensitas dan kualitas spektral, berfungsi sebagai sinyal morfogenik yang signifikan dalam sistem kultur jaringan tanaman. Penelitian komposisi spektral telah mengungkapkan respons yang berbeda tergantung pada panjang gelombang, dengan cahaya biru umumnya mendorong organogenesis tunas sementara panjang gelombang cahaya merah biasanya mendukung induksi akar. Paparan fotoperiodik berurutan—cahaya biru diikuti oleh cahaya merah—telah didokumentasikan secara efektif merangsang jalur organogenetik spesifik pada spesies tertentu.
Efek pengaturan dari panjang gelombang yang berbeda menunjukkan bagaimana kualitas cahaya dapat secara selektif mengendalikan hasil morfogenik. Pencahayaan fluoresen buatan menghasilkan respons yang bervariasi tergantung pada spesiesnya, mendorong pembentukan akar pada beberapa kultur sementara menghambatnya pada kultur lainnya. Beberapa spesies menunjukkan kebutuhan cahaya khusus, seperti yang diamati pada Pisum sativum (kacang polong), di mana inisiasi tunas pucuk terjadi secara optimal dalam kegelapan sebelum paparan cahaya merangsang perkembangan lebih lanjut.
Untuk sebagian besar penerapan kultur jaringan, protokol pencahayaan standar biasanya merekomendasikan iluminasi dengan intensitas sekitar 2.000-3.000 lux dengan periode fotoperiod 16 jam. Namun, spesies tertentu menunjukkan kebutuhan intensitas cahaya yang luar biasa, contohnya kultur kalus Nicotiana tabacum (tembakau), yang membutuhkan intensitas cahaya yang jauh lebih tinggi yaitu 10.000-15.000 lux untuk menginduksi pembentukan tunas pucuk atau embriogenesis somatik.
Suhu
Suhu berperan sebagai faktor lingkungan yang penting dalam sistem kultur jaringan tanaman, dengan suhu inkubasi optimal yang sangat bervariasi antar spesies berdasarkan kebutuhan habitat alaminya. Meskipun 25°C merupakan suhu inkubasi standar yang cocok untuk banyak spesies tanaman secara in vitro, adaptasi suhu spesifik spesies harus dipertimbangkan untuk memaksimalkan potensi organogenik. [
Spesies geofit dari daerah beriklim sedang biasanya membutuhkan rezim suhu yang lebih rendah daripada protokol standar. Contoh yang terkenal termasuk tanaman berumbi seperti Galanthus (snowdrop) yang menunjukkan pertumbuhan optimal pada suhu sekitar 15°C, sementara kultivar tertentu dari Narcissus (narsis) dan Allium (bawang hias) menunjukkan efisiensi regenerasi yang lebih baik pada suhu sekitar 18°C.
Sebaliknya, spesies asal tropis umumnya membutuhkan suhu yang lebih tinggi untuk pertumbuhan dan organogenesis optimal dalam kultur. Kultur kurma tumbuh subur pada suhu 27°C, sedangkan Monstera deliciosa (ceriman) menunjukkan kinerja regeneratif puncak pada suhu 30°C. Persyaratan suhu ini mencerminkan adaptasi evolusioner terhadap kondisi lingkungan asli tanaman tersebut.
Tingkat ploidi
Variasi jumlah kromosom, yaitu aneuploidi, poliploidi, dan lain-lain, dalam kultur sel tanaman telah didokumentasikan dengan baik di masa lalu. Ketidakstabilan kromosom sel mengakibatkan penurunan bertahap potensi morfogenetik jaringan kalus. Oleh karena itu, untuk mempertahankan potensi organogenik jaringan kalus dan stabilitas kromosom, disarankan agar waktu dan frekuensi subkultur dipantau secara teratur.
Zaman budaya
Usia kultur seringkali menjadi kunci keberhasilan organogenesis. Kultur muda/bahan yang baru dikultur ulang dapat menghasilkan organ lebih sering daripada kultur yang sudah tua. Alasan yang mungkin adalah penurunan atau hilangnya potensi organogenik pada kultur yang sudah tua. Namun, pada beberapa tanaman, kapasitas regenerasi tanaman dapat dipertahankan tanpa batas waktu selama bertahun-tahun.
Proses perkembangan
Dediferensiasi
Kemampuan sel untuk mengalami organogenesis sebagian besar bergantung pada penerapan zat pengatur pertumbuhan tanaman (PGR), yang memengaruhi arah perkembangan jaringan. Keseimbangan antara auksin dan sitokinin memainkan peran penting dalam menentukan apakah tunas atau akar akan terbentuk. Rasio auksin-sitokinin yang lebih rendah mendukung regenerasi tunas, sedangkan konsentrasi auksin yang lebih tinggi mendorong pembentukan akar. Misalnya, pada kultur Medicago sativa (alfalfa), peningkatan kadar kinetin yang dikombinasikan dengan konsentrasi 2,4-D (auksin sintetis) yang rendah menyebabkan perkembangan tunas, sedangkan peningkatan 2,4-D sambil mengurangi konsentrasi kinetin mendorong pembentukan akar. Namun, keberhasilan organogenesis tidak hanya bergantung pada perlakuan PGR. Ukuran fisik kalus atau jaringan yang berkembang harus mencapai ambang batas tertentu untuk mendukung pembentukan organ yang tepat, yang menyoroti pentingnya pensinyalan antar sel dalam menyelaraskan proses perkembangan.
Induksi
Fase induksi dalam organogenesis mewakili periode transisi antara jaringan yang mencapai kompetensi dan menjadi sepenuhnya siap untuk memulai pembentukan primordia. Selama tahap ini, jalur genetik terintegrasi mengarahkan proses perkembangan sebelum diferensiasi morfologis terjadi. Penelitian menunjukkan bahwa faktor kimia dan fisik tertentu dapat mengganggu jalur perkembangan yang diprogram secara genetik, mengubah hasil morfogenik. Dalam kasus Convolvulus arvensis, pengaruh eksternal ini ditemukan menghambat pembentukan tunas, dan malah mengarah pada perkembangan kalus.
Tahap induksi ditandai dengan sel atau kelompok sel yang berkomitmen untuk membentuk tunas atau akar. Penentuan ini diuji dengan memindahkan jaringan dari media yang diberi zat pengatur pertumbuhan ke media dasar yang mengandung mineral esensial, vitamin, dan sumber karbon tetapi tanpa zat pengatur pertumbuhan tanaman. Pada tahap ini, jaringan menyelesaikan proses induksi dan sepenuhnya menentukan nasib perkembangannya
Konsep kunci dalam proses ini adalah kanalisasi, yang mengacu pada kemampuan jalur perkembangan untuk secara konsisten menghasilkan fenotipe standar meskipun terdapat potensi variasi genetik atau lingkungan. Jika eksplan dikeluarkan dari media penginduksi tunas sebelum kanalisasi penuh terjadi, pembentukan tunas akan berkurang secara signifikan, dan perkembangan akar menjadi hasil yang dominan. Fenomena ini menyoroti plastisitas morfogenik jaringan tanaman secara in vitro, menunjukkan kemampuan mereka untuk menyesuaikan diri dengan kondisi eksternal dan isyarat perkembangan.
Diferensiasi
Selama fase ini, proses diferensiasi morfologis dimulai, yang mengarah pada pembentukan dan perkembangan organ yang baru terbentuk. Inisiasi organogenesis ditandai dengan pergeseran polaritas yang jelas, diikuti oleh pembentukan simetri radial dan pertumbuhan selanjutnya di sepanjang sumbu yang baru ditentukan, yang pada akhirnya membentuk tonjolan struktural yang menandai inisiasi organ.
Perkembangan organogenesis secara berurutan dapat diamati pada spesies seperti Pinus oocarpa Schiede, di mana tunas pucuk diregenerasi langsung dari kotiledon melalui organogenesis langsung. Namun, pola perkembangan spesifik dapat bervariasi di berbagai spesies tanaman yang ditanam secara in vitro. Perkembangan pembentukan organ meliputi perubahan morfologi yang berbeda, dimulai dengan perubahan tekstur permukaan, munculnya meristemoid, dan perluasan daerah meristematik baik secara vertikal maupun horizontal. Hal ini diikuti oleh penonjolan daerah meristematik di luar lapisan epidermis, pembentukan meristem terstruktur dengan primordia daun yang terlihat, dan akhirnya, perkembangan penuh tunas adventif.
Ciri khas kultur organogenik in vitro adalah pembentukan simultan beberapa meristemoid pada satu eksplan, dengan berbagai tingkat diferensiasi. Dalam eksplan yang sama, tunas dapat berada dalam berbagai tahap perkembangan, mulai dari inisiasi awal hingga struktur yang sepenuhnya berkembang. Setelah tunas memanjang melebihi panjang 1 cm, tunas tersebut dipindahkan ke substrat perakaran in vitro atau ex vitro, memungkinkan penyelesaian regenerasi plantlet dan pembentukan tanaman yang sepenuhnya terbentuk.
Keuntungan dan keterbatasan
Dalam proses organogenesis langsung, tunas aksiler dihasilkan langsung dari meristem yang sudah ada sebelumnya yang terletak di ujung tunas dan buku, sehingga menawarkan tingkat perbanyakan yang tinggi. Salah satu keunggulan utama metode ini adalah kemungkinan terjadinya mutasi yang rendah pada meristem tunas yang terorganisir, memastikan bahwa tanaman yang dihasilkan mempertahankan konsistensi genetik. Teknik ini sangat berharga untuk produksi dan konservasi tanaman yang penting secara ekonomi dan lingkungan, karena memungkinkan pembentukan banyak tunas secara efisien dari satu eksplan, menjaga keseragaman di seluruh tanaman yang diperbanyak. Lebih lanjut, semua tanaman yang dihasilkan melalui organogenesis langsung adalah tanaman asli, artinya mereka adalah klon genetik dari tanaman aslinya. [ diperlukan kutipan ]
Namun, terdapat beberapa keterbatasan pada organogenesis. Variasi somaklonal, yang dapat mengakibatkan keragaman genetik yang tidak diinginkan, merupakan masalah potensial, terutama dalam proses organogenesis tidak langsung. Selain itu, teknik ini mungkin tidak cocok untuk spesies tanaman yang sulit dikultur, yaitu spesies yang tidak merespon dengan baik terhadap kultur in vitro atau protokol regenerasi. Keterbatasan ini menyoroti perlunya penelitian berkelanjutan dan optimasi metode untuk berbagai spesies tanaman guna mengatasi tantangan ini dalam perbanyakan dan konservasi tanaman .
Pemanjangan sel
Selain pertumbuhan melalui pembelahan sel, tumbuhan juga dapat tumbuh melalui pemanjangan sel . Hal ini terjadi ketika sel-sel individual atau kelompok sel tumbuh lebih panjang. Tidak semua sel tumbuhan tumbuh dengan panjang yang sama. Ketika sel-sel di satu sisi batang tumbuh lebih panjang dan lebih cepat daripada sel-sel di sisi lainnya, batang akan membengkok ke sisi sel yang tumbuh lebih lambat. Pertumbuhan terarah ini dapat terjadi melalui respons tumbuhan terhadap rangsangan tertentu, seperti cahaya ( fototropisme ), gravitasi ( gravitropisme ), air ( hidrotropisme ), dan kontak fisik ( tigmotropisme ).


Pertumbuhan dan perkembangan tanaman dimediasi oleh hormon tanaman spesifik dan regulator pertumbuhan tanaman (PGR) (Ross et al. 1983). Tingkat hormon endogen dipengaruhi oleh umur tanaman, ketahanan terhadap dingin, dormansi, dan kondisi metabolisme lainnya; fotoperiod, kelembapan, suhu, dan kondisi lingkungan eksternal lainnya; dan sumber PGR eksogen, misalnya, yang diaplikasikan secara eksternal dan berasal dari rizosfer
Variasi morfologi selama pertumbuhan
Tumbuhan menunjukkan variasi alami dalam bentuk dan strukturnya. Meskipun semua organisme bervariasi dari individu ke individu, tumbuhan menunjukkan jenis variasi tambahan. Dalam satu individu, bagian-bagian tertentu berulang yang mungkin berbeda bentuk dan strukturnya dari bagian serupa lainnya. Variasi ini paling mudah dilihat pada daun tumbuhan, meskipun organ lain seperti batang dan bunga juga dapat menunjukkan variasi serupa. Ada tiga penyebab utama variasi ini: efek posisi, efek lingkungan, dan masa muda.

Terdapat variasi di antara bagian-bagian tanaman dewasa yang dihasilkan dari posisi relatif tempat organ tersebut diproduksi. Misalnya, di sepanjang cabang baru, daun-daun dapat bervariasi dalam pola yang konsisten di sepanjang cabang tersebut. Bentuk daun yang dihasilkan di dekat pangkal cabang berbeda dengan daun yang dihasilkan di ujung tanaman, dan perbedaan ini konsisten dari cabang ke cabang pada tanaman tertentu dan dalam spesies tertentu.
Cara struktur baru matang saat diproduksi dapat dipengaruhi oleh titik dalam siklus hidup tanaman ketika struktur tersebut mulai berkembang, serta oleh lingkungan tempat struktur tersebut terpapar. Suhu memiliki banyak pengaruh pada tanaman tergantung pada berbagai faktor, termasuk ukuran dan kondisi tanaman serta suhu dan durasi paparan. Semakin kecil dan semakin lunak tanaman, semakin besar kerentanannya terhadap kerusakan atau kematian akibat suhu yang terlalu tinggi atau terlalu rendah. Suhu memengaruhi laju proses biokimia dan fisiologis, laju yang umumnya (dalam batas tertentu) meningkat seiring dengan peningkatan suhu.
Juvenilitas atau heteroblasti adalah kondisi ketika organ dan jaringan yang dihasilkan oleh tanaman muda, seperti redih, seringkali berbeda dari organ dan jaringan yang dihasilkan oleh tanaman yang sama ketika sudah dewasa. Misalnya, pohon muda akan menghasilkan cabang yang lebih panjang dan ramping yang tumbuh ke atas lebih banyak daripada cabang yang akan dihasilkannya sebagai pohon dewasa. Selain itu, daun yang dihasilkan selama pertumbuhan awal cenderung lebih besar, lebih tipis, dan lebih tidak beraturan daripada daun pada tanaman dewasa. Spesimen tanaman muda mungkin terlihat sangat berbeda dari tanaman dewasa dari spesies yang sama sehingga serangga yang bertelur tidak mengenali tanaman tersebut sebagai makanan untuk anak-anak mereka. Transisi dari bentuk pertumbuhan awal ke akhir kadang-kadang disebut perubahan fase vegetatif . [10]
Struktur adventif
Struktur tumbuhan, termasuk akar, tunas, dan pucuk, yang berkembang di lokasi yang tidak biasa disebut adventif .
Akar dan tunas adventif biasanya berkembang di dekat jaringan vaskular yang ada sehingga dapat terhubung ke pembuluh kayu dan pembuluh tapis . Namun, lokasi pastinya sangat bervariasi. Pada batang muda, akar adventif sering terbentuk dari parenkim di antara berkas vaskular . Pada batang dengan pertumbuhan sekunder, akar adventif sering berasal dari parenkim floem di dekat kambium vaskular . Pada stek batang, akar adventif kadang-kadang juga berasal dari sel kalus yang terbentuk di permukaan potongan. Stek daun Crassula membentuk akar adventif di epidermis.
Tunas dan pucuk
Tunas adventif berkembang dari tempat selain meristem apikal pucuk, yang terdapat di ujung batang, atau pada buku pucuk, di ketiak daun, tunas tersebut dibiarkan di sana selama pertumbuhan primer. Tunas adventif dapat berkembang pada akar atau daun, atau pada pucuk sebagai pertumbuhan baru. Meristem apikal pucuk menghasilkan satu atau lebih tunas aksiler atau lateral pada setiap buku. Ketika batang menghasilkan pertumbuhan sekunder yang cukup besar, tunas aksiler dapat hancur. Tunas adventif kemudian dapat berkembang pada batang dengan pertumbuhan sekunder
Tunas adventif sering terbentuk setelah batang terluka atau dipangkas . Tunas adventif membantu menggantikan cabang yang hilang. Tunas dan tunas adventif juga dapat berkembang pada batang pohon dewasa ketika batang yang teduh terkena sinar matahari yang terang karena pohon-pohon di sekitarnya ditebang. Pohon ki merah ( Sequoia sempervirens ) sering mengembangkan banyak tunas adventif di bagian bawah batangnya. Jika batang utama mati, batang baru sering tumbuh dari salah satu tunas adventif. Potongan kecil batang ki merah dijual sebagai suvenir yang disebut bungkong ki merah. Potongan-potongan ini ditempatkan dalam wadah berisi air, dan tunas adventif akan tumbuh membentuk tunas.
Beberapa tumbuhan biasanya mengembangkan tunas adventif pada akarnya, yang dapat memanjang cukup jauh dari tumbuhan. Tunas yang tumbuh dari tunas adventif pada akar disebut tunas liar . Ini adalah jenis reproduksi vegetatif alami pada banyak spesies, misalnya banyak rumput, Populus tremuloides, dan waga-waga kanada . Pohon pando tumbuh dari satu batang menjadi 47.000 batang melalui pembentukan tunas adventif pada satu sistem akar. [ diperlukan kutipan ]
Beberapa daun mengembangkan tunas adventif, yang kemudian membentuk akar adventif, sebagai bagian dari reproduksi vegetatif ; misal Tolmiea menziesii dan Kalanchoe daigremontiana. Tunas adventif tersebut kemudian terlepas dari tanaman induk dan berkembang sebagai klon terpisah dari tanaman induk. [
Pentrubusan adalah praktik memotong batang pohon hingga ke tanah untuk mendorong pertumbuhan tunas adventif yang cepat. Secara tradisional, metode ini digunakan untuk menghasilkan tiang, bahan pagar, atau kayu bakar. Metode ini juga dipraktikkan untuk tanaman biomassa yang ditanam untuk bahan bakar, seperti pohon hawar atau dedalu
Akar

Perakaran adventif mungkin merupakan aklimatisasi penghindaran stres bagi beberapa spesies, yang dipicu oleh masukan seperti hipoksia [11] atau kekurangan nutrisi. Fungsi penting ekologis lainnya dari perakaran adventif adalah reproduksi vegetatif spesies pohon seperti Salix dan Sequoia di lingkungan tepi sungai . [12]
Kemampuan batang tanaman untuk membentuk akar adventif dimanfaatkan dalam perbanyakan komersial dengan stek . Pemahaman tentang mekanisme fisiologis di balik perakaran adventif telah memungkinkan beberapa kemajuan dalam meningkatkan perakaran stek dengan aplikasi auksin sintetis sebagai bubuk perakaran dan dengan penggunaan luka basal selektif. [13] Kemajuan lebih lanjut dapat dicapai di tahun-tahun mendatang dengan menerapkan penelitian tentang mekanisme pengaturan lainnya pada perbanyakan komersial dan dengan analisis komparatif kontrol molekuler dan ekofisiologis perakaran adventif pada spesies 'sulit berakar' vs. 'mudah berakar'.
Akar dan tunas adventif sangat penting ketika orang memperbanyak tanaman melalui stek, cangkok, dan kultur jaringan . Hormon tumbuhan, yang disebut auksin, sering diaplikasikan pada stek batang, pucuk, atau daun untuk mendorong pembentukan akar adventif, misalnya, daun violet afrika dan sedum serta pucuk kastuba dan miana . Perbanyakan melalui stek akar membutuhkan pembentukan tunas adventif, misalnya, pada lobak dan apel . Dalam cangkok, akar adventif terbentuk pada batang udara sebelum bagian batang dipotong untuk membuat tanaman baru. Tanaman hias besar sering diperbanyak dengan cangkok udara . Akar dan tunas adventif harus berkembang dalam perbanyakan tanaman melalui kultur jaringan.
Bentuk yang dimodifikasi
- Akar umbi tidak memiliki bentuk yang pasti; contoh: ubi jalar.
- Akar berfasikulasi (akar umbi) muncul dalam kelompok di pangkal batang; contoh: asparagus, dahlia.
- Akar nodulosa membengkak di dekat ujungnya; contoh: kunyit.
- Akar sangga muncul dari beberapa buku pertama batang. Akar ini menembus miring ke bawah ke dalam tanah dan memberikan dukungan pada tanaman; contoh: jagung, tebu.
- Akar penopang memberikan dukungan mekanis pada cabang udara. Cabang lateral tumbuh vertikal ke bawah ke dalam tanah dan bertindak sebagai pilar; contoh: beringin.
- Akar panjat yang muncul dari buku-buku tersebut menempel pada suatu penopang dan memanjatnya; contoh: sirih gading.
- Akar moniliform atau manik-manik akar yang berdaging memberikan tampilan seperti manik-manik, misalnya: pare, krokot.
Perkembangan daun
Genetika di balik perkembangan bentuk daun pada Arabidopsis thaliana telah dipecah menjadi tiga tahap: Inisiasi primordium daun, pembentukan dorsiventralitas, dan perkembangan meristem marginal. Primordium daun diinisiasi oleh penekanan gen dan protein dari famili KNOX kelas I (seperti SHOOT APICAL MERISTEMLESS ). Protein KNOX kelas I ini secara langsung menekan biosintesis gibberellin di primordium daun. Banyak faktor genetik ditemukan terlibat dalam penekanan gen-gen ini di primordium daun (seperti ASYMMETRIC LEAVES1, BLADE-ON-PETIOLE1, SAWTOOTH1, dll.). Dengan demikian, dengan penekanan ini, kadar gibberellin meningkat dan primordium daun memulai pertumbuhan.
Perkembangan bunga

Perkembangan bunga adalah proses di mana tumbuhan berbunga menghasilkan pola ekspresi gen di meristem yang mengarah pada munculnya organ yang berorientasi pada perkembangbiakan seksual, yaitu bunga . Ada tiga perkembangan fisiologis yang harus terjadi agar hal ini dapat berlangsung: pertama, tumbuhan harus beralih dari keadaan belum dewasa secara seksual ke keadaan dewasa secara seksual (yaitu transisi menuju pembungaan); kedua, transformasi fungsi meristem apikal dari meristem vegetatif menjadi meristem bunga atau perbungaan ; dan terakhir, pertumbuhan organ-organ bunga secara individual. Fase terakhir ini telah dimodelkan menggunakan model ABC, yang menggambarkan dasar biologis proses tersebut dari perspektif genetika molekuler dan perkembangan .
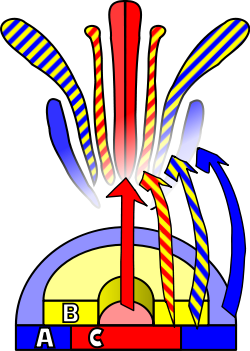
Diperlukan stimulus eksternal untuk memicu diferensiasi meristem menjadi meristem bunga. Stimulus ini akan mengaktifkan pembelahan sel mitosis di meristem, khususnya di sisi-sisinya tempat primordia baru terbentuk. Stimulus yang sama juga akan menyebabkan meristem mengikuti pola perkembangan yang akan mengarah pada pertumbuhan meristem bunga, berbeda dengan meristem vegetatif. Perbedaan utama antara kedua jenis meristem ini, selain perbedaan yang jelas antara organ target, adalah filotaksis vertisilat (atau melingkar), yaitu, tidak adanya pemanjangan batang di antara lingkaran atau vertisilat primordium yang berurutan. Vertikal ini mengikuti perkembangan akropetal, menghasilkan sepal, kelopak, benang sari, dan putik . Perbedaan lain dari meristem aksila vegetatif adalah bahwa meristem bunga bersifat "tertentu", yang berarti bahwa, setelah terdiferensiasi, sel-selnya tidak akan lagi membelah .
Identitas organ yang terdapat dalam empat vertisi bunga merupakan konsekuensi dari interaksi setidaknya tiga jenis produk gen, masing-masing dengan fungsi yang berbeda. Menurut model ABC, fungsi A dan C diperlukan untuk menentukan identitas vertisi perianth dan vertisi reproduktif. Fungsi-fungsi ini bersifat eksklusif dan ketiadaan salah satunya berarti bahwa yang lain akan menentukan identitas semua vertisi bunga. Fungsi B memungkinkan diferensiasi kelopak dari sepal pada vertisi sekunder, serta diferensiasi benang sari dari putik pada vertisi tersier.
Aroma bunga
Tumbuhan menggunakan bentuk bunga, bunga, dan aroma untuk menarik berbagai serangga untuk penyerbukan . Senyawa tertentu dalam aroma yang dipancarkan menarik penyerbuk tertentu. Pada Petunia hybrida, benzenoid volatil diproduksi untuk mengeluarkan aroma bunga. Meskipun komponen jalur biosintesis benzenoid telah diketahui, enzim dalam jalur tersebut, dan regulasi selanjutnya dari enzim tersebut, masih belum ditemukan. [14]
Untuk menentukan regulasi jalur, bunga P. hybrida Mitchell digunakan dalam mikrolarik spesifik kelopak untuk membandingkan bunga yang hampir menghasilkan aroma, dengan bunga kultivar P. hybrida W138 yang menghasilkan sedikit benzenoid volatil. cDNA gen dari kedua tanaman tersebut diurutkan. Hasilnya menunjukkan bahwa ada faktor transkripsi yang meningkat pada bunga Mitchell, tetapi tidak pada bunga W138 yang tidak memiliki aroma bunga. Gen ini diberi nama ODORANT1 (ODO1). Untuk menentukan ekspresi ODO1 sepanjang hari, dilakukan analisis gel sap RNA . Gel menunjukkan bahwa tingkat transkrip ODO1 mulai meningkat antara pukul 13.00 dan 16.00, mencapai puncaknya pada pukul 22.00 dan terendah pada pukul 10.00. Tingkat transkrip ODO1 ini secara langsung sesuai dengan garis waktu emisi benzenoid volatil. Selain itu, gel tersebut mendukung temuan sebelumnya bahwa bunga W138 yang tidak beraroma hanya memiliki sepersepuluh tingkat transkrip ODO1 dari bunga Mitchell. Dengan demikian, jumlah ODO1 yang dihasilkan sesuai dengan jumlah benzenoid volatil yang dikeluarkan, menunjukkan bahwa ODO1 mengatur biosintesis benzenoid. [15]
Gen tambahan yang bersumbangsing pada biosintesis senyawa aroma utama adalah OOMT1 dan OOMT2. OOMT1 dan OOMT2 membantu mensintesis orcinol O-metiltransferase (OOMT), yang mengkatalisis dua langkah terakhir jalur DMT, menghasilkan 3,5-dimetoksitoluena (DMT). DMT adalah senyawa aroma yang diproduksi oleh banyak jenis mawar, namun beberapa varietas mawar, seperti Rosa gallica dan mawar Damaskus Rosa damascene, tidak mengeluarkan DMT. Telah disarankan bahwa varietas ini tidak menghasilkan DMT karena mereka tidak memiliki gen OOMT. Namun, setelah percobaan imunolokalisasi, OOMT ditemukan di epidermis kelopak bunga. Untuk mempelajari hal ini lebih lanjut, kelopak bunga mawar disentrifugasi ultra . Supernatan dan pelet diperiksa dengan sap barat . Deteksi protein OOMT pada 150.000g dalam supernatan dan pelet memungkinkan para peneliti untuk menyimpulkan bahwa protein OOMT terkait erat dengan membran epidermis kelopak bunga. Percobaan tersebut menentukan bahwa gen OOMT memang ada di dalam varietas Rosa gallica dan mawar Damaskus Rosa damascene, namun gen OOMT tidak diekspresikan di jaringan bunga tempat DMT diproduksi. [16]
Rujukan
- ^ Bäurle, I; Laux, T (2003). "Apical meristems: The plant's fountain of youth". BioEssays. 25 (10): 961–70. doi:10.1002/bies.10341. PMID 14505363. Review.
- ^ Drost, Hajk-Georg; Janitza, Philipp; Grosse, Ivo; Quint, Marcel (2017). "Cross-kingdom comparison of the developmental hourglass". Current Opinion in Genetics & Development. 45: 69–75. doi:10.1016/j.gde.2017.03.003. PMID 28347942.
- ^ Irie, Naoki; Kuratani, Shigeru (2011-03-22). "Comparative transcriptome analysis reveals vertebrate phylotypic period during organogenesis". Nature Communications (dalam bahasa Inggris). 2. Bibcode:2011NatCo...2..248I. doi:10.1038/ncomms1248. ISSN 2041-1723. PMC 3109953. PMID 21427719.
- ^ Domazet-Lošo, Tomislav; Tautz, Diethard (2010-12-09). "A phylogenetically based transcriptome age index mirrors ontogenetic divergence patterns". Nature (dalam bahasa Inggris). 468 (7325): 815–818. Bibcode:2010Natur.468..815D. doi:10.1038/nature09632. ISSN 0028-0836. PMID 21150997.
- ^ Quint, Marcel; Drost, Hajk-Georg; Gabel, Alexander; Ullrich, Kristian Karsten; Bönn, Markus; Grosse, Ivo (2012-10-04). "A transcriptomic hourglass in plant embryogenesis". Nature (dalam bahasa Inggris). 490 (7418): 98–101. Bibcode:2012Natur.490...98Q. doi:10.1038/nature11394. ISSN 0028-0836. PMID 22951968.
- ^ Drost, Hajk-Georg; Gabel, Alexander; Grosse, Ivo; Quint, Marcel (2015-05-01). "Evidence for Active Maintenance of Phylotranscriptomic Hourglass Patterns in Animal and Plant Embryogenesis". Molecular Biology and Evolution (dalam bahasa Inggris). 32 (5): 1221–1231. doi:10.1093/molbev/msv012. ISSN 0737-4038. PMC 4408408. PMID 25631928.
- ^ Leopold, A. Carl (1964). animal and there young one. McGraw-Hill. hlm. 183.
- ^ Brand, U; Hobe, M; Simon, R (2001). "Functional domains in plant shoot meristems". BioEssays. 23 (2): 134–41. doi:10.1002/1521-1878(200102)23:2<134::AID-BIES1020>3.0.CO;2-3. PMID 11169586. Review.
- ^ Barlow, P (2005). "Patterned cell determination in a plant tissue: The secondary phloem of trees". BioEssays. 27 (5): 533–41. doi:10.1002/bies.20214. PMID 15832381.
- ^ Jones, Cynthia S. (1999-11-01). "An Essay on Juvenility, Phase Change, and Heteroblasty in Seed Plants". International Journal of Plant Sciences. 160 (S6): 105–S111. Bibcode:1999IJPlS.160S.105J. doi:10.1086/314215. ISSN 1058-5893. PMID 10572025.
- ^ Drew, M. C.; Jackson, M. B.; Giffard, S. (1979). "Ethylene-promoted adventitious rooting and development of cortical air spaces (Aerenchyma) in roots may be adaptive responses to flooding in Zea mays L". Planta. 147 (1): 83–88. Bibcode:1979Plant.147...83D. doi:10.1007/BF00384595. PMID 24310899.
- ^ Naiman, Robert J.; Decamps, Henri (1997). "The Ecology of Interfaces: Riparian Zones". Annual Review of Ecology and Systematics. 28 (1): 621–658. Bibcode:1997AnRES..28..621N. doi:10.1146/annurev.ecolsys.28.1.621. JSTOR 2952507.
- ^ De Klerk, Geert-Jan; Van Der Krieken, Wim; De Jong, Joke C. (1999). "Review the formation of adventitious roots: New concepts, new possibilities". In Vitro Cellular & Developmental Biology - Plant. 35 (3): 189–199. Bibcode:1999IVCDB..35..189D. doi:10.1007/s11627-999-0076-z.
- ^ Schuurink, Robert C.; Haring, Michel A.; Clark, David G. (2006). "Regulation of volatile benzenoid biosynthesis in petunia flowers". Trends in Plant Science. 11 (1): 20–25. doi:10.1016/j.tplants.2005.09.009. PMID 16226052.
- ^ Schuurink, Robert C.; Haring, Michel A.; Clark, David G. (2006). "Regulation of volatile benzenoid biosynthesis in petunia flowers". Trends in Plant Science. 11 (1): 20–25. doi:10.1016/j.tplants.2005.09.009. PMID 16226052.
- ^ Scalliet, Gabriel; Lionnet, Claire; Le Bechec, Mickaël; Dutron, Laurence; Magnard, Jean-Louis; Baudino, Sylvie; Bergougnoux, Véronique; Jullien, Frédéric; Chambrier, Pierre (2006-01-01). "Role of Petal-Specific Orcinol O -Methyltransferases in the Evolution of Rose Scent". Plant Physiology. 140 (1): 18–29. doi:10.1104/pp.105.070961. ISSN 1532-2548. PMC 1326028. PMID 16361520.
Konten ini disalin dari wikipedia, mohon digunakan dengan bijak.

